A cikk orvosi szakértője

Új kiadványok
Petefészek
Utolsó ellenőrzés: 04.07.2025

Minden iLive-tartalmat orvosi szempontból felülvizsgáltak vagy tényszerűen ellenőriznek, hogy a lehető legtöbb tényszerű pontosságot biztosítsák.
Szigorú beszerzési iránymutatásunk van, és csak a jó hírű média oldalakhoz, az akadémiai kutatóintézetekhez és, ha lehetséges, orvosilag felülvizsgált tanulmányokhoz kapcsolódik. Ne feledje, hogy a zárójelben ([1], [2] stb.) Szereplő számok ezekre a tanulmányokra kattintható linkek.
Ha úgy érzi, hogy a tartalom bármely pontatlan, elavult vagy más módon megkérdőjelezhető, jelölje ki, és nyomja meg a Ctrl + Enter billentyűt.
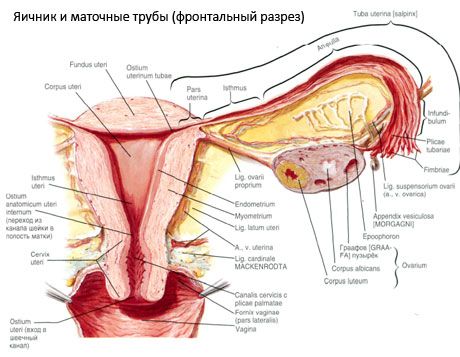
A petefészek (ovarium; görögül oophoron) egy páros szerv, női ivarmirigy, amely a medenceüregben található, a méh széles szalagja mögött. A petefészkekben fejlődnek és érnek a női ivarsejtek (petesejtek), és képződnek a női nemi hormonok, amelyek a vérbe és a nyirokba jutnak. A petefészek ovális alakú, anteroposterior irányban kissé lapított. A petefészek színe rózsaszínes. A szült nő petefészkének felszínén bemélyedések és hegek láthatók - az ovuláció és a sárgatestek átalakulásának nyomai. A petefészek súlya 5-8 g. A petefészek méretei: hossza 2,5-5,5 cm, szélessége 1,5-3,0 cm, vastagsága - legfeljebb 2 cm. A petefészeknek két szabad felszíne van: a középső felszín (facies medialis), amely a medenceüreg felé néz, és amelyet részben a petevezeték borít, valamint az oldalsó felszín (facies lateralis), amely a medence oldalfalához közel, egy enyhén kifejezett bemélyedéshez - a petefészek árokához - csatlakozik. Ez a fossa a felül hashártyával borított külső csípőerek, alul pedig a méh- és obturátor artériák közötti szögben található. A petefészek mögött a megfelelő oldali húgyvezeték retroperitoneálisan halad felülről lefelé.
A petefészek felszíne egy domború szabad (hátsó) szélbe (margo liber), elöl pedig a bélfodri szélbe (margo mesovaricus) torkollik, amely a hashártya (petefészek bélfodra) rövid redőjével kapcsolódik a méh széles szalagjának hátsó levélkéjéhez. A szerv ezen elülső szélén egy barázdált mélyedés található - a petefészek hilumja (hilum ovarii), amelyen keresztül az artéria és az idegek belépnek a petefészekbe, a vénák és a nyirokerek pedig kilépnek. A petefészeknek is két vége van: a lekerekített felső csőszerű vége (extremitas tubaria), amely a petevezeték felé néz, és az alsó méhszerű vége (extremitas utenna), amelyet a petefészek szalagja (lig. ovarii proprium) köt a méhhez. Ez a szalag, körülbelül 6 mm vastag kerek zsinór formájában, a petefészek méh felőli végétől a méh oldalsó szögletéig terjed, amely a méh széles szalagjának két levele között helyezkedik el. A petefészek szalagrendszeréhez tartozik még a petefészek szalag-szuszpenzoriuma (lig.suspensorium ovarii), amely a hashártya redője, amely a kis medence falától a petefészekig fut, és benne a petefészek ereit és rostos rostkötegeit tartalmazza. A petefészket egy rövid bélfodor (mesovarium) rögzíti, amely a hashártya megkettőződése, amely a méh széles szalagjának hátsó levelétől a petefészek bélfodros széléig fut. Magukat a petefészkeket nem fedi a hashártya. A petevezeték legnagyobb petefészek-pereme a petefészek csőszerű végéhez kapcsolódik. A petefészek topográfiája a méh helyzetétől, méretétől (terhesség alatt) függ. A petefészkek a kismedencei üreg nagyon mozgékony szervei.
 [
[ A petefészek erei és idegei
A petefészkek vérellátását az aa. et vv. ovaricae et uterinae biztosítja. Mindkét petefészek artéria (aa. ovaricae dextra et sinistra) az aorta elülső felszínéről, közvetlenül a veseartériák alatt ered; a jobb oldali általában az aortából, a bal oldali pedig a veseartériából ered. A nagy horpaszizmok elülső felszíne mentén lefelé és oldalirányban haladva mindkét petefészek artéria keresztezi az elülső húgyvezetéket (ágakat adva hozzá), a külső csípőereket, a határvonalat, és belép a medenceüregbe, itt a petefészek felfüggesztő szalagjában található. Mediális irányban haladva a petefészek artéria a méh széles szalagjának levelei között áthalad a petevezeték alatt, ágakat adva hozzá, majd a petefészek bélfodrozódik; belép a petefészek hilumába.
A petefészek artéria ágai széles körben anasztomózist alkotnak a méhartéria petefészek ágaival. A petefészkekből a vénás kiáramlás elsősorban a petefészek vénás plexusába történik, amely a petefészek-hilum régiójában található. Innen a vér kiáramlása két irányban történik: a méh- és a petefészek vénáin keresztül. A jobb petefészek vénája billentyűkkel rendelkezik, és az alsó üres vénába ömlik. A bal petefészek vénája a bal vesevénába ömlik, bár nincsenek billentyűi.
A petefészkekből a nyirokelvezetés a nyirokereken keresztül történik, különösen bőségesen a szervkapu területén, ahol a szubovariális nyirokfonat kitűnik. Ezután a nyirok a petefészek nyirokereinek mentén a paraaortikus nyirokcsomókba ürül.
A petefészkek beidegzése
Szimpatikus - a lisztérzékenység (szoláris), a mesenterialis felső és a hypogastricus plexusok posztganglionáris rostjai biztosítják; paraszimpatikus - a zsigeri szakrális idegek biztosítják.
A petefészek szerkezete
A petefészek felszínét egyrétegű germinális hám borítja. Alatta sűrű kötőszöveti fehérjeburok (tunica albuginea) fekszik. A petefészek kötőszövete alkotja a stromáját (stroma ovarii), amely rugalmas rostokban gazdag. A petefészek anyaga, parenchymája külső és belső rétegre oszlik. A petefészek közepén, a kapujához közelebb található belső réteget velőnek (medulla ovarii) nevezik. Ebben a rétegben, a laza kötőszövetben számos vér- és nyirokér, valamint ideg található. A petefészek külső rétege - a kéreg (cortex ovarii) sűrűbb. Sok kötőszövetet tartalmaz, amelyben az érő primer petefészek tüszők (folliculi ovarici primarii), a másodlagos (vezikuláris) tüszők (folliculi ovarici secundarii, s.vesiculosi), valamint az érett tüszők, a Graaf-tüszők (folliculi ovarici maturis), valamint a sárga- és atretikus testek találhatók.
Minden tüsző egy női reproduktív petesejtet, vagy oocitát (ovocytus) tartalmaz. A petesejt átmérője legfeljebb 150 µm, kerek, sejtmagot és nagy mennyiségű citoplazmát tartalmaz, amely a sejtszervecskék mellett fehérje-lipid zárványokat (sárgája) és glikogént is tartalmaz, amelyek a petesejt táplálásához szükségesek. A petesejt általában az ovulációt követő 12-24 órán belül felhasználja a tápanyagkészletét. Ha a megtermékenyítés nem történik meg, a petesejt elpusztul.
Az emberi petesejtet két membrán borítja. Belül található a citolemma, ami a petesejt citoplazmatikus membránja. A citolemmán kívül úgynevezett tüszősejtek rétege található, amelyek védik a petesejtet és hormonképző funkciót látnak el - ösztrogént választanak ki.
A méh, a petevezetékek és a petefészkek fiziológiai helyzetét a hashártyát, a szalagokat és a medence szövetét egyesítő felfüggesztő, rögzítő és támasztó apparátusok biztosítják. A felfüggesztő apparátust páros képződmények képviselik, magukban foglalják a méh kerek és széles szalagjait, a megfelelő szalagokat és a petefészkek felfüggesztő szalagjait. A méh széles szalagjai, a petefészkek megfelelő és felfüggesztő szalagjai tartják a méhet középső helyzetben. A kerek szalagok előrehúzzák a méhfenéket, és biztosítják annak fiziológiai dőlését.
A rögzítő (horgonyzó) szerkezet biztosítja a méh helyzetét a kismedence közepén, és gyakorlatilag lehetetlenné teszi az oldalirányú, előre-hátra történő elmozdulását. Mivel azonban a szalagszerkezet a méh alsó szakaszától eltávolodik, a méh különböző irányú dőlése lehetséges. A rögzítő szerkezet magában foglalja a medence laza szövetében elhelyezkedő szalagokat, amelyek a méh alsó szakaszától a medence oldalsó, elülső és hátsó faláig nyúlnak: a keresztcsonti, a kardinális, a méh-vezikális és a hólyagos-szeméremcsonti szalagokat.
A mesovarium mellett a következő petefészek szalagokat különböztetjük meg:
- A petefészek felfüggesztő szalagja, korábban infundibulpelvicus szalagnak nevezték. A hashártya redője, amelyen áthaladnak a petefészek erei (a. et v. ovarica), nyirokerei és idegei, és amely a medence oldalfala, az ágyéki fascia (a közös csípőartéria külső és belső részre osztódásának területén) és a petefészek felső (petevezeték) vége között húzódik;
- A petefészek megfelelő szalagja a széles méhszalag levelei között halad, közelebb a hátsó levélhez, és a petefészek alsó végét köti össze a méh oldalsó szélével. A petefészek megfelelő szalagja a petevezeték kezdete és a kerek szalag között, az utóbbi mögött és felett kapcsolódik a méhhez. A szalag vastagságában haladnak az ováriumok, amelyek a méhartéria végágai;
- vakbél-petefészek szalag A szalag a vakbél bélfodrozódásától a méh jobb petefészkéig vagy széles szalagjáig terjed, hashártya-redő formájában. A szalag inkonzisztens, és a nők 1/2-1/3-ánál figyelhető meg.
A tartószerkezetet a medencefenék izmai és fasciája képviseli, amelyek alsó, középső és felső (belső) rétegekre oszlanak.
A legerősebb a felső (belső) izomréteg, amelyet a végbélnyílást emelő páros izom képvisel. Izomkötegekből áll, amelyek a farkcsonttól a medencecsontokig három irányban terjednek szét (pubococcygeus, iliococcygeus és ischiococcygeus). Ezt az izomréteget medencei rekeszizomnak is nevezik.
A középső izomréteg a szimfízis, a szeméremcsont és az ülőcsont között helyezkedik el. A középső izomréteg - az urogenitális rekeszizom - a medence kijáratának elülső felét foglalja el, amelyen keresztül a húgycső és a hüvely áthalad. A lemezei között elülső részben izomkötegek találhatók, amelyek a húgycső külső záróizmát alkotják, hátsó részben pedig izomkötegek, amelyek haránt irányban haladnak - a gát mély harántizma.
A medencefenék izmainak alsó (külső) rétege felületes izmokból áll, amelyek alakja a 8-as számra hasonlít. Ezek közé tartozik a bulbocavernous, az ischiocavernous, a külső anális záróizom és a felületes haránt gátizmok.
A petefészkek ontogenezise
A tüszőnövekedés és atresia folyamata a terhesség 20. hetében kezdődik, és a szülés idejére akár 2 millió petesejt is marad a lány petefészkeiben. A menarche-ra számuk 300 ezerre csökken. A reproduktív élet teljes időszaka alatt legfeljebb 500 tüsző éri el az érettséget és ovulál. A tüszők kezdeti növekedése nem függ az FSH stimulációjától, korlátozott, és az atresia gyorsan bekövetkezik. Úgy vélik, hogy a szteroid hormonok helyett a helyi autokrin/parakrin peptidek a primer tüszők növekedésének és atresiájának fő szabályozói. Úgy tartják, hogy a tüszőnövekedés és az atresia folyamatát semmilyen fiziológiai folyamat nem szakítja félbe. Ez a folyamat minden életkorban folytatódik, beleértve a méhen belüli időszakot és a menopauzát is, és a terhesség, az ovuláció és az anovuláció szakítja félbe. A tüszők növekedését és számát kiváltó mechanizmus az egyes ciklusokban még mindig nem tisztázott.
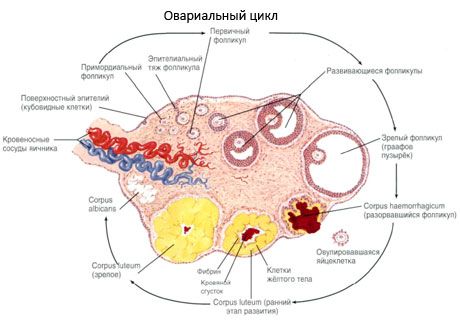
Fejlődése során a tüsző több fejlődési szakaszon megy keresztül. Az őscsírasejtek a petezsák endodermájából, az allantoisból származnak, és a terhesség 5-6. hetében az embrió nemi szervi területére vándorolnak. A gyors mitotikus osztódás eredményeként, amely a terhesség 6-8. hetétől a 16-20. hetéig tart, az embrió petefészkeiben akár 6-7 millió petesejt is képződik, amelyeket egy vékony granulosa sejtréteg vesz körül.
Preantrális tüsző - a petesejtet membrán (Zona pellucida) veszi körül. A petesejtet körülvevő granulosa sejtek proliferációnak indulnak, növekedésük a gonadotropinoktól függ és korrelál az ösztrogénszinttel. A granulosa sejtek az FSH célpontjai. A preantrális tüsző stádiumában a granulosa sejtek háromféle szteroid szintetizálására képesek: túlnyomórészt az aromatáz aktivitását indukálja, amely a fő enzim, amely az androgéneket ösztradiollá alakítja. Úgy vélik, hogy az ösztradiol képes növelni saját receptorainak számát, közvetlen mitogén hatást gyakorolva a granulosa sejtekre az FSH-tól függetlenül. Parakrin faktornak tekintik, amely fokozza az FSH hatásait, beleértve az aromatizációs folyamatok aktiválását is.
Az FSH receptorok a granulosa sejtek membránjain jelennek meg, amint a tüsző növekedése megkezdődik. Az FSH csökkenése vagy növekedése a receptorok számának megváltozásához vezet. Az FSH ezen hatását növekedési faktorok modulálják. Az FSH a G-proteinen, az adenilát-cikláz rendszeren keresztül hat, bár a tüszőben a szteroidogenezist főként az FSH szabályozza, ebben a folyamatban számos tényező vesz részt: ioncsatornák, tirozin-kináz receptorok, a másodlagos hírvivők foszfolipáz rendszere.
Az androgének szerepe a korai tüszőfejlődésben összetett. A granulosa sejtek androgénreceptorokkal rendelkeznek. Ezek nemcsak szubsztrátjai az FSH által kiváltott ösztrogénekké történő aromatizációnak, hanem alacsony koncentrációban is fokozhatják az aromatizációt. Amikor az androgénszint megemelkedik, a preantrális granulosa sejtek előnyben részesítik az ösztrogének aromatizációs útvonalát, hanem az egyszerűbb androgén útvonalat az 5a-reduktázon keresztül, hogy olyan androgénné alakuljanak, amely nem alakítható át ösztrogénné, ezáltal gátolva az aromatáz aktivitását. Ez a folyamat gátolja az FSH és LH receptorok képződését is, így megállítva a tüszőfejlődést.
Az aromatizációs folyamat során a magas androgénszintű tüsző atresián megy keresztül. A tüsző növekedése és fejlődése attól függ, hogy képes-e az androgéneket ösztrogénekké alakítani.
FSH jelenlétében a tüszőfolyadék domináns anyaga az ösztrogének lesznek. FSH hiányában az androgének. Az LH normális esetben a ciklus közepéig hiányzik a tüszőfolyadékból. Amint az LH-szint emelkedik, a granulosa sejtek mitotikus aktivitása csökken, degeneratív elváltozások jelennek meg, és a tüszőben megnő az androgénszint. A tüszőfolyadékban a szteroidok szintje magasabb, mint a plazmában, és a petefészek sejtek: a granulosa és a theca sejtek funkcionális aktivitását tükrözi. Ha az FSH egyetlen célpontja a granulosa sejtek, akkor az LH-nak sok célpontja van - a theca sejtek, a stromális és a lutein sejtek, valamint a granulosa sejtek. Mind a granulosa, mind a theca sejtek képesek szteroidogenezisre, de az aromatáz aktivitás dominál a granulosa sejtekben.
Az LH-ra válaszul a theca sejtek androgéneket termelnek, amelyeket aztán a granulosa sejtek FSH által indukált aromatizáció révén ösztrogénekké alakítanak.
Ahogy a tüsző növekszik, a theca sejtek elkezdik expresszálni az LH receptorok, a P450 sec és a 3béta-hidroxiszteroid dehidrogenáz génjeit, az inzulinszerű növekedési faktor (IGF-1) szinergikusan az LH-val növeli a génexpressziót, de nem stimulálja a szteroidogenezist.
A petefészek szteroidogenezise mindig LH-függő. Ahogy a tüsző növekszik, a theca sejtek expresszálják a P450c17 enzimet, amely koleszterinből androgént képez. A granulosa sejtek nem rendelkeznek ezzel az enzimmel, és a theca sejtektől függenek, hogy ösztrogéneket termeljenek androgénekből. A szteroidogenezissel ellentétben a follikulogenezis FSH-függő. Ahogy a tüsző növekszik és az ösztrogénszint emelkedik, egy visszacsatolási mechanizmus aktiválódik - az FSH-termelés gátolt, ami viszont a tüsző aromatáz aktivitásának csökkenéséhez, és végső soron apoptózison (programozott sejthalálon) keresztül tüszőatresiához vezet.
Az ösztrogének és az FSH visszacsatolási mechanizmusa gátolja a növekedésnek indult tüszők fejlődését, de nem a domináns tüszőét. A domináns tüsző több FSH-receptort tartalmaz, amelyek támogatják a granulosa sejtek proliferációját és az androgének ösztrogénekké aromatizálódását. Ezenkívül a parakrin és autokrin útvonalak fontos koordinátorként működnek az antrális tüszőfejlődésben.
Az autokrin/parakrin szabályozó peptidekből (inhibin, aktivin, follisztatin) áll, amelyeket a granulosa sejtek szintetizálnak FSH-ra válaszul, és bejutnak a tüszőfolyadékba. Az inhibin csökkenti az FSH szekréciót; az aktivin serkenti az FSH felszabadulását az agyalapi mirigyből és fokozza az FSH hatását a petefészekben; a follisztatin gátolja az FSH aktivitását, valószínűleg az aktinhoz való kötődés révén. Az ovuláció és a sárgatest kifejlődése után az inhibin az LH szabályozása alatt áll.
A petefészeksejtek növekedését és differenciálódását az inzulinszerű növekedési faktorok (IGE) befolyásolják. Az IGF-1 a granulosa sejtekre hat, ami a ciklikus adenozin-monofoszfát (cAMP), a progeszteron, az oxitocin, a proteoglikán és az inhibin szintjének növekedését okozza.
Az IGF-1 a theca sejtekre hat, fokozott androgéntermelést okozva. A theca sejtek viszont tumor nekrózis faktort (TNF) és epidermális növekedési faktort (EGF) termelnek, amelyeket szintén az FSH szabályoz.
Az EGF serkenti a granulosa sejtek proliferációját. Az IGF-2 a tüszőfolyadék fő növekedési faktora, és az IGF-1, a TNF-α, a TNF-3 és az EGF is megtalálható benne.
A petefészekműködés parakrin és/vagy autokrin szabályozásának zavara szerepet játszik az ovulációs zavarokban és a policisztás petefészkek kialakulásában.
Ahogy az antrális tüsző növekszik, a tüszőfolyadék ösztrogéntartalma növekszik. Növekedésük csúcspontján LH-receptorok jelennek meg a granulosa sejteken, a granulosa sejtek luteinizációja megtörténik, és a progeszterontermelés fokozódik. Így a preovulációs időszakban az ösztrogéntermelés fokozódása LH-receptorok megjelenését okozza, az LH pedig a granulosa sejtek luteinizációját és a progeszterontermelést okozza. A progeszteronszint növekedése csökkenti az ösztrogénszintet, ami látszólag az FSH második csúcsát okozza a ciklus közepén.
Az ovuláció az LH-csúcs után 10-12 órával, az ösztradiol-csúcs után pedig 24-36 órával következik be. Az LH-ról úgy tartják, hogy serkenti a petesejt redukciós osztódását, a granulosa sejtek sárgatestképződését, valamint a progeszteron és a prosztaglandin szintézisét a tüszőben.
A progeszteron fokozza a proteolitikus enzimek aktivitását, amelyek a prosztaglandinnal együtt részt vesznek a tüszőfal megrepedésében. A progeszteron által kiváltott FSH csúcs lehetővé teszi, hogy a petesejt a plazminogént a plazmin proteolitikus enzimmé alakítva kilépjen a tüszőből, és elegendő számú LH-receptort biztosít a luteális fázis normális fejlődéséhez.
Az ovulációt követő 3 napon belül a granulosa sejtek mérete megnő, és jellegzetes, lutein pigmenttel töltött vakuólumok jelennek meg bennük. A theca-luteális sejtek differenciálódnak a thecától és a stromától, és a sárgatest részévé válnak. A sárgatestbe behatoló kapillárisok az angiogenezis faktorok hatására nagyon gyorsan fejlődnek, és a javuló vaszkularizációval fokozódik a progeszteron és az ösztrogének termelése. A szteroidogenezis aktivitását és a sárgatest élettartamát az LH szintje határozza meg. A sárgatest nem homogén sejtes képződmény. A 2 típusú sárgatestsejt mellett endothelsejteket, makrofágokat, fibroblasztokat stb. tartalmaz. A nagy sárgatestsejtek peptideket (relaxin, oxitocin) termelnek, és aktívabbak a szteroidogenezisben, nagyobb aromatáz aktivitással és nagyobb progeszteronszintézissel, mint a kis sejtek.
A progeszteron csúcsa az LH-csúcs utáni 8. napon jelentkezik. Megjegyzendő, hogy a progeszteron és az ösztradiol epizodikusan választódik ki a luteális fázisban, az LH pulzáló kibocsátásával korrelációban. A sárgatest kialakulásával az inhibin termelésének szabályozása az FSH-ról az LH-ra kerül át. Az inhibin szintje az LH-csúcs előtti ösztradiolszint-növekedéssel együtt növekszik, és az LH-csúcs után is tovább növekszik, bár az ösztrogénszint csökken. Bár az inhibint és az ösztradiolt a granulosa sejtek választják ki, szabályozásuk különböző útvonalakon keresztül történik. Az inhibin szintjének csökkenése a luteális fázis végén hozzájárul az FSH szintjének emelkedéséhez a következő ciklusban.
A sárgatest nagyon gyorsan csökken - az ovuláció utáni 9-11. napon.
A degeneráció mechanizmusa nem tisztázott, és nem kapcsolódik az ösztrogének luteolitikus szerepéhez vagy egy receptorhoz kapcsolódó mechanizmushoz, ahogyan azt az endometriumban megfigyelték. Létezik egy másik magyarázat is a sárgatest által termelt ösztrogének szerepére. Ismert, hogy az ösztrogének szükségesek a progeszteronreceptorok szintéziséhez az endometriumban. A sárgatesti fázisban lévő ösztrogének valószínűleg szükségesek a progeszteronnal összefüggő változásokhoz az endometriumban az ovuláció után. A progeszteronreceptorok nem megfelelő fejlődése, az elégtelen ösztrogénszint következtében, valószínűleg további mechanizmus a meddőséghez és a korai vetélésekhez, ami a sárgatesti fázis hiányának egy másik formája. Úgy vélik, hogy a sárgatest élettartama az ovuláció idején alakul ki. És minden bizonnyal visszafejlődik, ha a terhesség miatt nem támogatja a humán koriongonadotropin. Így a sárgatest regressziója az ösztradiol, a progeszteron és az inhibin szintjének csökkenéséhez vezet. Az inhibin csökkenése megszünteti az FSH-ra gyakorolt elnyomó hatását; Az ösztradiol és a progeszteron szintjének csökkenése lehetővé teszi a GnRH-szekréció nagyon gyors helyreállását és a visszacsatolási mechanizmus megszűnését az agyalapi mirigyből. Az inhibin és az ösztradiol szintjének csökkenése, valamint a GnRH szintjének emelkedése az FSH termelődését eredményezi az LH helyett. Az FSH szintjének emelkedése tüszőnövekedéshez vezet, majd egy domináns tüsző kiválasztásához, és új ciklus kezdődik, ha a terhesség nem következik be. A szteroid hormonok vezető szerepet játszanak a reprodukciós biológiában és az általános fiziológiában. Meghatározzák az emberi fenotípust, befolyásolják a szív- és érrendszert, a csontok anyagcseréjét, a bőrt, az általános jólétet, és kulcsszerepet játszanak a terhességben. A szteroid hormonok hatása tükrözi azokat az intracelluláris és genetikai mechanizmusokat, amelyek szükségesek ahhoz, hogy egy extracelluláris jel juttasson a sejtmagba, és fiziológiai választ váltson ki.
Az ösztrogének diffúz módon behatolnak a sejtmembránba, és a sejtmagban található receptorokhoz kötődnek. A receptor-szteroid komplex ezután a DNS-hez kötődik. A célsejtekben ezek a kölcsönhatások génexpresszióhoz, fehérjeszintézishez, valamint specifikus sejt- és szövetfunkcióhoz vezetnek.