A cikk orvosi szakértője

Új kiadványok
agyalapi mirigy
Utolsó ellenőrzés: 04.07.2025

Minden iLive-tartalmat orvosi szempontból felülvizsgáltak vagy tényszerűen ellenőriznek, hogy a lehető legtöbb tényszerű pontosságot biztosítsák.
Szigorú beszerzési iránymutatásunk van, és csak a jó hírű média oldalakhoz, az akadémiai kutatóintézetekhez és, ha lehetséges, orvosilag felülvizsgált tanulmányokhoz kapcsolódik. Ne feledje, hogy a zárójelben ([1], [2] stb.) Szereplő számok ezekre a tanulmányokra kattintható linkek.
Ha úgy érzi, hogy a tartalom bármely pontatlan, elavult vagy más módon megkérdőjelezhető, jelölje ki, és nyomja meg a Ctrl + Enter billentyűt.
Az agyalapi mirigy (hypophysis, s.glandula pituitaria) az ékcsont tátongó üregének (sella turcica) agyalapi mirigyében található, és az agy dura mater nyúlványa választja el a koponyaüregtől, alkotva a sella rekeszizmát (sella rekeszizma). Ezen a rekeszizom nyílásán keresztül kapcsolódik az agyalapi mirigy a diencephalon hipotalamuszának infundibulumához. Az agyalapi mirigy haránt mérete 10-17 mm, az anteroposterior 5-15 mm, a függőleges 5-10 mm. Az agyalapi mirigy tömege férfiakban körülbelül 0,5 g, nőkben 0,6 g. Az agyalapi mirigyet kívülről egy tok borítja.
Az agyalapi mirigy két különböző rudimentből való fejlődésének megfelelően a szervben két lebeny különböztethető meg - az elülső és a hátsó. Az adenohypophysis vagy elülső lebeny (adenohypophysis, s.lobus anterior) nagyobb, az agyalapi mirigy teljes tömegének 70-80%-át teszi ki. Sűrűbb, mint a hátsó lebeny. Az elülső lebenyben megkülönböztethető a disztális rész (pars distalis), amely az agyalapi mirigy árok elülső részét foglalja el, egy közbenső rész (pars intermedia), amely a hátsó lebeny határán helyezkedik el, és egy gumós rész (pars tuberalis), amely felfelé halad és a hipotalamusz infundibulumához kapcsolódik. A vérerek bősége miatt az elülső lebeny halványsárga színű, vöröses árnyalattal. Az elülső agyalapi mirigy parenchymáját többféle mirigysejt képviseli, amelyek szálai között sinusoidális vérkapillárisok helyezkednek el. Az adenohipofízis sejtek fele (50%) kromofil adenociták, amelyek citoplazmájában finomszemcsés szemcsék találhatók, amelyek jól festődnek krómsókkal. Ezek az acidofil adenociták (az összes adenohipofízis sejt 40%-a) és a bazofil adenociták (10%). A bazofil adenociták közé tartoznak a gonadotrop, kortikotrop és tireotrop endokrinociták. A kromofób adenociták kicsik, nagy maggal és kis mennyiségű citoplazmával rendelkeznek. Ezeket a sejteket a kromofil adenociták prekurzorainak tekintik. Az adenohipofízis sejtek másik 50%-a kromofób adenocita.
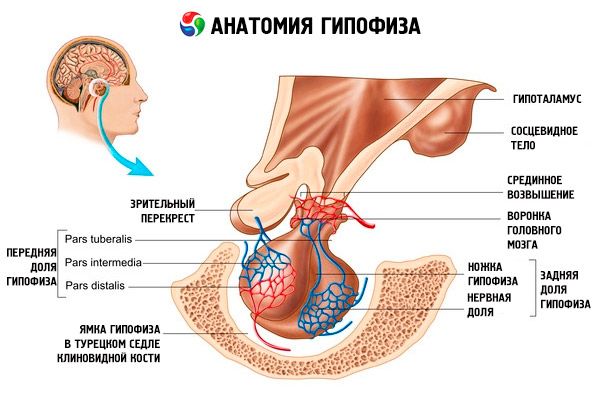
A neurohypofízis, vagy hátsó lebeny (neurohypophysis, s.lobus posterior) az agyalapi mirigy árok hátsó részében található ideglebenyből (lobus nervosus) és az adenohypophízis gumós része mögött elhelyezkedő tölcsérből (infundibulum) áll. Az agyalapi mirigy hátsó lebenyét neurogliasejtek (agyalapi mirigy sejtek), a hipotalamusz neuroszekréciós magvaiból a neurohypophízisbe tartó idegrostok és neuroszekréciós részecskék alkotják.
Az agyalapi mirigy idegrostok (pályák) és vérerek útján funkcionálisan kapcsolódik a diencephalon hipotalamuszához, amely az agyalapi mirigy működését szabályozza. Az agyalapi mirigyet és a hipotalamuszt neuroendokrin, érrendszeri és idegi kapcsolataikkal együtt általában a hipotalamusz-hipofízis rendszernek tekintik.
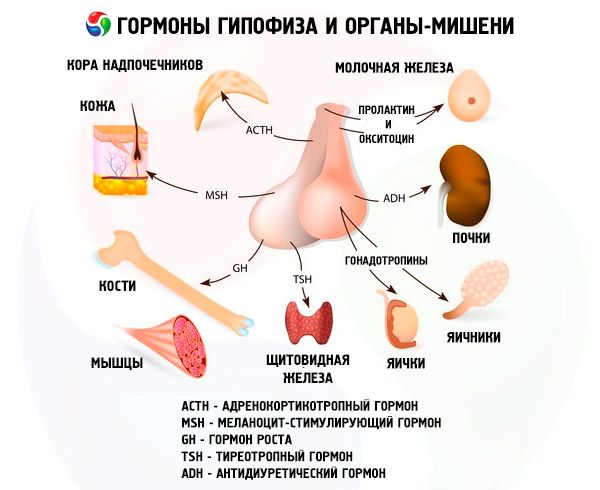
Az elülső és hátsó agyalapi mirigy hormonjai a szervezet számos funkcióját befolyásolják, elsősorban más endokrin mirigyeken keresztül. Az elülső agyalapi mirigyben az acidofil adenociták (alfa-sejtek) szomatotrop hormont (növekedési hormont) termelnek, amely részt vesz a fiatal szervezetek növekedési és fejlődési folyamatainak szabályozásában. A kortikotrop endokrinociták adrenokortikotrop hormont (ACTH) választanak ki, amely serkenti a mellékvesék általi szteroid hormonok szekrécióját. A tireotrop endokrinociták tireotrop hormont (TSH) választanak ki, amely befolyásolja a pajzsmirigy fejlődését és aktiválja hormonjainak termelését. A gonadotrop hormonok: a tüszőstimuláló (FSH), a luteinizáló (LH) és a prolaktin - befolyásolják a szervezet szexuális érését, szabályozzák és serkentik a tüszők fejlődését a petefészekben, az ovulációt, az emlőmirigyek növekedését és a tejtermelést nőknél, a spermatogenezis folyamatát férfiaknál. Ezeket a hormonokat bazofil adenociták (béta-sejtek) termelik. Az agyalapi mirigy lipotrop faktorai is itt választódnak ki, amelyek befolyásolják a zsírok mobilizálását és hasznosulását a szervezetben. Az elülső lebeny köztes részében melanocita-stimuláló hormon képződik, amely a pigmentek - melaninok - képződését szabályozza a szervezetben.
A hipotalamuszban található supraoptikus és paraventrikuláris magok neuroszekréciós sejtjei vazopresszint és oxitocint termelnek. Ezek a hormonok a hipotalamusz-hipofízis traktust alkotó axonok mentén a hátsó agyalapi mirigy sejtjeibe jutnak. A hátsó agyalapi mirigyből ezek az anyagok a véráramba kerülnek. A vazopresszin hormon érszűkítő és antidiuretikus hatású, ezért antidiuretikus hormonnak (ADH) is nevezik. Az oxitocin stimuláló hatással van a méhizmok összehúzódására, fokozza a tejelválasztást a tejelválasztást a tejeltávolító emlőmirigyben, gátolja a sárgatest fejlődését és működését, valamint befolyásolja a gyomor-bél traktus simaizmainak (nem harántcsíkolt) tónusának változásait.
Az agyalapi mirigy fejlődése
Az agyalapi mirigy elülső lebenye a szájüreg hátsó falának hámjából fejlődik ki gyűrű alakú kinövésként (Rathke-tasak). Ez az ektodermális nyúlvány a leendő harmadik kamra alja felé növekszik. Felé, a második agyhólyag (a leendő harmadik kamra alja) alsó felszínéről egy nyúlvány nő, amelyből az infundibulum szürke tuberkuluma és az agyalapi mirigy hátsó lebenye fejlődik ki.
Az agyalapi mirigy erei és idegei
A belső nyaki verőerekből és az agy artériás körének ereiből a felső és az alsó agyalapi mirigy artériái az agyalapi mirigybe vezetnek. A felső agyalapi mirigy artériák a szürke magba és a hipotalamusz infundibulumba vezetnek, itt egymással anasztomózist képeznek, és kapillárisokat képeznek, amelyek áthatolnak az agyszöveten - az elsődleges hemokapilláris hálózaton. Ennek a hálózatnak a hosszú és rövid hurkaiból képződnek a portális vénák, amelyek az agyalapi mirigy elülső lebenyébe vezetnek. Az agyalapi mirigy elülső lebenyének parenchymájában ezek a vénák széles sinusoidális kapillárisokra bomlanak, kialakítva a másodlagos hemokapilláris hálózatot. Az agyalapi mirigy hátsó lebenyét főként az alsó agyalapi mirigy artéria látja el vérrel. A felső és az alsó agyalapi mirigy artériák között hosszú artériás anasztomózisok vannak. A vénás vér kiáramlása a másodlagos hemokapilláris hálózatból egy vénás rendszeren keresztül történik, amely az agy dura materének kavernózus és interkavernózus sinusaiba áramlik.
Az agyalapi mirigy beidegzésében részt vesznek a szimpatikus rostok, amelyek az artériákkal együtt jutnak be a szervbe. A posztganglionáris szimpatikus idegrostok a belső carotis artéria plexusából indulnak ki. Ezenkívül az agyalapi mirigy hátsó lebenyében számos, a hipotalamusz magjaiban található neuroszekréciós sejtek nyúlványainak vége található.
Az agyalapi mirigy életkorral összefüggő jellemzői
Az agyalapi mirigy átlagos súlya újszülötteknél eléri a 0,12 g-ot. A szerv súlya 10 éves korra megduplázódik, 15 éves korra pedig megháromszorozódik. 20 éves korra az agyalapi mirigy súlya eléri a maximumát (530-560 mg), és a következő életkorokban szinte változatlan marad. 60 év után a belső elválasztású mirigy súlyának enyhe csökkenése figyelhető meg.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Agyalapi mirigy hormonjai
Az idegi és hormonális szabályozás egységét a szervezetben az agyalapi mirigy és a hipotalamusz szoros anatómiai és funkcionális kapcsolata biztosítja. Ez a komplexum határozza meg az egész endokrin rendszer állapotát és működését.
Az agyalapi mirigy a fő endokrin mirigy, amely számos peptidhormont termel, amelyek közvetlenül szabályozzák a perifériás mirigyek működését. Vöröses-szürke, bab alakú képződmény, amelyet 0,5-0,6 g súlyú rostos tok borít. Mérete kissé változik az egyén nemétől és életkorától függően. Az agyalapi mirigy általánosan elfogadott felosztása két lebenyre maradt, amelyek fejlettségükben, szerkezetükben és működésükben különböznek: az elülső disztális - adenohipofízisre és a hátsó - neurohipofízisre. Az első a mirigy teljes tömegének körülbelül 70%-át teszi ki, és hagyományosan disztális, infundibuláris és közbenső részre, a második pedig hátsó részre vagy lebenyre és az agyalapi mirigy nyélére oszlik. A mirigy az ékcsont turkálójának agyalapi mirigyárkában található, és a nyélen keresztül kapcsolódik az agyhoz. Az elülső lebeny felső részét a chiasma látóideg és a látóidegpályák borítják. Az agyalapi mirigy vérellátása igen bőséges, és a belső carotis artéria ágain (a felső és alsó agyalapi mirigy artériákon), valamint az agy artériás körének ágain keresztül történik. A felső agyalapi mirigy artériák az adenohypophysis, az alsóbbak pedig a neurohypophysis vérellátásában vesznek részt, érintkezve a hipotalamusz nagysejtes magjainak axonjainak neurosecretory végződéseivel. Az előbbiek a hipotalamusz eminentia medianusába jutnak, ahol kapilláris hálózatba (az elsődleges kapilláris plexusba) szétszóródva. Ezek a kapillárisok (amellyel a mediobasalis hipotalamusz kis neurosecretory sejtjeinek axonjainak végződései érintkeznek) portális vénákban gyűlnek össze, amelyek az agyalapi mirigy nyélén lefelé haladnak az adenohypophysis parenchymájába, ahol ismét sinusoidális kapillárisok hálózatává válnak (a másodlagos kapilláris plexusba). Így a vér, miután előzőleg áthaladt a hipotalamusz eminentia medianusán, ahol hipotalamusz adenohypophysotrop hormonokkal (felszabadító hormonokkal) dúsul, az adenohypophysisbe jut.
A másodlagos plexus számos kapillárisából az adenohypophysealis hormonokkal telített vér kiáramlása a vénák rendszerén keresztül történik, amelyek viszont a dura mater vénás sinusaiba, majd az általános véráramba áramlanak. Így az agyalapi mirigy portális rendszere, amelynek véráramlása a hipotalamuszból csökkenő irányban történik, az adenohypophysis tropikus funkcióinak neurohumorális szabályozásának komplex mechanizmusának morfofunkcionális komponense.
Az agyalapi mirigyet szimpatikus rostok idegzik be, amelyek az agyalapi mirigy artériáit követik. Ezek a posztganglionáris rostokból erednek, amelyek a carotis internán haladnak át, és a felső nyaki ganglionokhoz kapcsolódnak. Az adenohypophysisnek nincs közvetlen beidegzése a hipotalamuszból. A hátsó lebeny a hipotalamusz neurosecretory magvaiból kapja az idegrostokat.
Az adenohipofízis szövettani felépítését tekintve nagyon összetett képződmény. Kétféle mirigysejt található benne - kromofób és kromofil. Ez utóbbiak viszont acidofil és bazofil sejtekre oszlanak (az agyalapi mirigy részletes szövettani leírása a kézikönyv megfelelő részében található). Meg kell azonban jegyezni, hogy az adenohipofízis parenchymáját alkotó mirigysejtek által termelt hormonok - az utóbbiak sokfélesége miatt - kémiai jellegükben bizonyos mértékig eltérőek, és a kiválasztó sejtek finomszerkezetének meg kell felelnie mindegyikük bioszintézisének jellemzőinek. De néha megfigyelhetők az adenohipofízisben a mirigysejtek átmeneti formái, amelyek képesek több hormon termelésére. Bizonyítékok vannak arra, hogy az adenohipofízis mirigysejtjeinek típusa nem mindig genetikailag meghatározott.
A sella turcica rekeszizom alatt található az elülső lebeny tölcsér alakú része. Ez átöleli az agyalapi mirigy nyélét, és érintkezik a szürke tuberkulusszal. Az adenohypophysis ezen részét hámsejtek jelenléte és bőséges vérellátása jellemzi. Hormonálisan is aktív.
Az agyalapi mirigy közbenső (középső) része több réteg nagy szekréciós-aktív bazofil sejtből áll.
Az agyalapi mirigy hormonjain keresztül különféle funkciókat lát el. Elülső lebenye adrenokortikotrop (ACTH), pajzsmirigy-stimuláló (TSH), tüszőstimuláló (FSH), sárgatestképző (LH), lipotrop hormonokat, valamint növekedési hormont - szomatotrop (STO) és prolaktint termel. A középső lebenyben a melanocita-stimuláló hormon (MSH) szintetizálódik, a hátsó lebenyben pedig a vazopresszin és az oxitocin halmozódik fel.
ACTH
Az agyalapi mirigy hormonjai fehérje- és peptidhormonok, valamint glikoproteinek csoportját alkotják. Az elülső agyalapi mirigy hormonjai közül az ACTH-t tanulmányozták a legjobban. Bazofil sejtek termelik. Fő fiziológiai funkciója a szteroid hormonok bioszintézisének és szekréciójának serkentése a mellékvesekéreg által. Az ACTH melanocita-stimuláló és lipotrop aktivitást is mutat. 1953-ban tiszta formában izolálták. Később megállapították kémiai szerkezetét, amely 39 aminosavmaradékból áll emberben és számos emlősben. Az ACTH-nak nincs fajspecificitása. Jelenleg mind magának a hormonnak, mind molekulájának különböző, a természetes hormonoknál aktívabb fragmenseinek kémiai szintézise folyik. A hormon szerkezete a peptidlánc két szakaszából áll, amelyek közül az egyik biztosítja az ACTH receptorhoz való kötődését és kimutatását, a másik pedig biológiai hatást fejt ki. Úgy tűnik, hogy a hormon és a receptor elektromos töltéseinek kölcsönhatásán keresztül kötődik az ACTH receptorhoz. Az ACTH biológiai effektorának szerepét a 4-10 molekula fragmentuma (Met-Glu-His-Phen-Arg-Tri-Tri) tölti be.
Az ACTH melanocita-stimuláló aktivitása a molekulában található N-terminális régiónak köszönhető, amely 13 aminosavmaradékból áll, és az alfa-melanocita-stimuláló hormon szerkezetét ismétli. Ez a régió egy heptapeptidet is tartalmaz, amely más agyalapi mirigy hormonokban is megtalálható, és adrenokortikotrop, melanocita-stimuláló és lipotrop aktivitással rendelkezik.
Az ACTH hatásának kulcsmomentuma a protein-kináz enzim citoplazmában történő aktiválódása a cAMP részvételével. A foszforilált protein-kináz aktiválja az észteráz enzimet, amely a koleszterin-észtereket szabad anyaggá alakítja a zsírcseppekben. A riboszómák foszforilációjának eredményeként a citoplazmában szintetizált fehérje serkenti a szabad koleszterin kötődését a citokróm P-450-hez, és annak lipidcseppekből a mitokondriumokba történő átjutását, ahol minden olyan enzim jelen van, amely biztosítja a koleszterin kortikoszteroidokká történő átalakulását.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Pajzsmirigy-stimuláló hormon
A TSH - tireotropin - a pajzsmirigy fejlődésének és működésének, a pajzsmirigyhormonok szintézisének és szekréciójának fő szabályozója. Ez az összetett fehérje - glikoprotein - alfa és béta alegységekből áll. Az első alegység szerkezete megegyezik a luteinizáló hormon alfa alegységével. Sőt, nagyrészt megegyezik a különböző állatfajokban. Az emberi TSH béta alegységében található aminosav-maradékok szekvenciája megfejtve 119 aminosav-maradékból áll. Megjegyezhető, hogy az emberi és a szarvasmarha TSH béta alegységei sok tekintetben hasonlóak. A glikoprotein hormonok biológiai tulajdonságait és biológiai aktivitásának jellegét a béta alegység határozza meg. Ez biztosítja a hormon kölcsönhatását a különböző célszervek receptoraival is. A legtöbb állatban a béta alegység azonban csak azután mutat specifikus aktivitást, hogy egyesül az alfa alegységgel, amely a hormon egyfajta aktivátoraként működik. Ez utóbbi azonos valószínűséggel indukál luteinizáló, tüszőstimuláló és tireotrop aktivitást, amelyet a béta alegység tulajdonságai határoznak meg. A talált hasonlóság arra enged következtetni, hogy ezek a hormonok egy közös elődből fejlődtek ki az evolúció során, a béta alegység határozza meg a hormonok immunológiai tulajdonságait is. Feltételezhető, hogy az alfa alegység védi a béta alegységet a proteolitikus enzimek hatásától, és megkönnyíti annak szállítását az agyalapi mirigyből a perifériás célszervekhez.
Gonadotrop hormonok
A gonadotropinok LH és FSH formájában vannak jelen a szervezetben. Ezen hormonok funkcionális célja általában a reprodukciós folyamatok biztosítására korlátozódik mindkét nemnél. A TSH-hoz hasonlóan összetett fehérjék - glikoproteinek. Az FSH a női petefészkekben a tüszők érését idézi elő, a férfiaknál pedig serkenti a spermatogenezist. Az LH a nőknél tüszőrepedést okoz a sárgatest kialakulásával, és serkenti az ösztrogének és a progeszteron szekrécióját. Férfiaknál ugyanaz a hormon gyorsítja az intersticiális szövet fejlődését és az androgének szekrécióját. A gonadotropinok hatásai egymástól függenek és szinkronban jelentkeznek.
A gonadotropin-szekréció dinamikája nőknél a menstruációs ciklus során változik, és kellő részletességgel tanulmányozták. A ciklus preovulációs (follikuláris) fázisában az LH-tartalom meglehetősen alacsony szinten van, míg az FSH-szint megemelkedik. A tüsző érésével az ösztradiol-szekréció fokozódik, ami elősegíti az agyalapi mirigy fokozott gonadotropin-termelését, valamint mind az LH-, mind az FSH-ciklusok bekövetkezését, azaz a nemi szteroidok serkentik a gonadotropinok szekrécióját.
Jelenleg az LH szerkezete ismert. A TSH-hoz hasonlóan 2 alegységből áll: a-ból és b-ből. Az LH alfa alegységének szerkezete a különböző állatfajokban nagyrészt megegyezik, megfelel a TSH alfa alegységének szerkezetének.
Az LH béta alegységének szerkezete jelentősen eltér a TSH béta alegységének szerkezetétől, bár négy azonos, 4-5 aminosavmaradékból álló peptidlánc-szakasszal rendelkezik. A TSH-ban ezek a 27-31., 51-54., 65-68. és 78-83. pozíciókban lokalizálódnak. Mivel az LH és a TSH béta alegysége határozza meg a hormonok specifikus biológiai aktivitását, feltételezhető, hogy az LH és a TSH szerkezetében lévő homológ szakaszoknak biztosítaniuk kell a béta alegységek kapcsolódását az alfa alegységhez, és a szerkezetben eltérő szakaszoknak kell felelősnek lenniük a hormonok biológiai aktivitásának specificitásáért.
A natív LH nagyon stabil a proteolitikus enzimek hatásával szemben, de a béta alegységet a kimotripszin gyorsan hasítja, az α alegységet pedig az enzim nehezen hidrolizálja, azaz védő szerepet tölt be, megakadályozza, hogy a kimotripszin hozzáférjen a peptidkötésekhez.
Ami az FSH kémiai szerkezetét illeti, a kutatók még nem jutottak végleges eredményekre. Az LH-hoz hasonlóan az FSH is két alegységből áll, de az FSH béta alegysége eltér az LH béta alegységétől.
Prolaktin
Egy másik hormon, a prolaktin (laktogén hormon) aktív szerepet játszik a szaporodási folyamatokban. Az emlősökben a prolaktin fő fiziológiai tulajdonságai az emlőmirigyek és a laktáció fejlődésének stimulálásában, a faggyúmirigyek és a belső szervek növekedésében nyilvánulnak meg. Elősegíti a szteroidok másodlagos nemi jellegekre gyakorolt hatásának megnyilvánulását hímekben, serkenti a sárgatest szekréciós aktivitását egerekben és patkányokban, és részt vesz a zsíranyagcsere szabályozásában. Az utóbbi években nagy figyelmet fordítottak a prolaktinra, mint az anyai viselkedés szabályozójára; ezt a polifunkcionalitást evolúciós fejlődése magyarázza. Ez az ősi agyalapi mirigy hormonok egyike, és még a kétéltűekben is megtalálható. Jelenleg a prolaktin szerkezetét egyes emlősfajokban teljesen megfejtették. A tudósok azonban egészen a közelmúltig kétségeiket fejezték ki egy ilyen hormon létezésével kapcsolatban az emberekben. Sokan úgy vélték, hogy funkcióját a növekedési hormon látja el. Most meggyőző bizonyítékokat szereztek a prolaktin jelenlétéről az emberekben, és szerkezetét részben megfejtették. A prolaktin receptorok aktívan kötődnek a növekedési hormonhoz és a placenta laktogénjéhez, ami a három hormon egyetlen hatásmechanizmusára utal.
Szomatotropin
A növekedési hormon, a szomatotropin, még szélesebb hatásspektrummal rendelkezik, mint a prolaktin. A prolaktinhoz hasonlóan az adenohipofízis acidofil sejtjei termelik. Az STH serkenti a csontváz növekedését, aktiválja a fehérjebioszintézist, zsírmobilizáló hatással rendelkezik, és elősegíti a testméret növekedését. Ezenkívül koordinálja az anyagcsere-folyamatokat.
A hormon utóbbiban való részvételét megerősíti az agyalapi mirigy szekréciójának hirtelen növekedése, például amikor a vércukorszint csökken.
Ennek az emberi hormonnak a kémiai szerkezete mára teljesen ismert – 191 aminosavmaradék. Elsődleges szerkezete hasonló a koriongyökérhez vagy a placenta-laktogénhez. Ezek az adatok a két hormon jelentős evolúciós közelségére utalnak, bár biológiai aktivitásukban eltéréseket mutatnak.
Szükséges hangsúlyozni a szóban forgó hormon magas fajspecificitását - például az állati eredetű STH inaktív az emberben. Ezt mind az emberi és állati STH receptorok közötti reakció, mind maga a hormon szerkezete magyarázza. Jelenleg folynak vizsgálatok az STH komplex szerkezetében található aktív központok azonosítására, amelyek biológiai aktivitást mutatnak. A molekula olyan egyedi fragmenseit is vizsgálják, amelyek más tulajdonságokat mutatnak. Például az emberi STH pepszinnel történő hidrolízise után izoláltak egy 14 aminosavmaradékból álló peptidet, amely a 31-44. molekulaszakasznak felel meg. Nem volt növekedési hatása, de lipotrop aktivitásban jelentősen meghaladta a natív hormont. Az emberi növekedési hormon, ellentétben az állatok hasonló hormonjával, jelentős laktogén aktivitással rendelkezik.
Az adenohipofízis számos peptid- és fehérjeanyagot szintetizál, amelyek zsírmobilizáló hatással rendelkeznek, az agyalapi mirigy tropikus hormonjai - ACTH, STH, TSH és mások - pedig lipotrop hatásúak. Az utóbbi években különösen a béta- és γ-lipotrop hormonok (LPG) kaptak különös hangsúlyt. A béta-LPG biológiai tulajdonságait vizsgálták a legrészletesebben; a lipotrop aktivitás mellett melanocita-stimuláló, kortikotropin-stimuláló és hipokalcémiás hatással is rendelkezik, valamint inzulinszerű hatást is kivált.
Jelenleg a juh LPG (90 aminosavmaradék), a sertések és szarvasmarhák lipotrop hormonjainak elsődleges szerkezetét megfejtették. Ez a hormon fajspecifikus, bár a béta-LPG központi régiójának szerkezete a különböző fajokban azonos. Ez határozza meg a hormon biológiai tulajdonságait. E régió egyik fragmense megtalálható az alfa-MSH, a béta-MSH, az ACTH és a béta-LPG szerkezetében. Felmerült, hogy ezek a hormonok ugyanabból az előanyagból származnak az evolúció során. A γ-LPG gyengébb lipotrop aktivitással rendelkezik, mint a béta-LPG.
Melanocita-stimuláló hormon
Ez a hormon, amelyet az agyalapi mirigy középső lebenyében szintetizálnak, biológiai funkciójában serkenti a melanin bőrpigment bioszintézisét, elősegíti a kétéltűek bőrében a pigmentsejtek, a melanociták méretének és számának növekedését. Az MSH ezen tulajdonságait a hormon biológiai vizsgálatában használják fel. A hormonnak két típusa van: alfa- és béta-MSH. Kimutatták, hogy az alfa-MSH nem rendelkezik fajspecifikussággal, és minden emlősben azonos kémiai szerkezettel rendelkezik. Molekulája egy 13 aminosavmaradékból álló peptidlánc. A béta-MSH ezzel szemben fajspecifikus, és szerkezete a különböző állatokban eltérő. A legtöbb emlősben a béta-MSH molekula 18 aminosavmaradékból áll, és csak az emberben hosszabbodik meg az aminosav végétől négy aminosavmaradékkal. Meg kell jegyezni, hogy az alfa-MSH rendelkezik némi adrenokortikotrop aktivitással, és hatása az állatok és az emberek viselkedésére ma már bizonyított.
Oxitocin és vazopresszin
A hipotalamuszban szintetizálódó vazopresszin és oxitocin az agyalapi mirigy hátsó lebenyében halmozódik fel: a vazopresszin a nucleus supraopticus neuronjaiban, az oxitocin pedig a nucleus paraventrikulatusban. Innen átkerülnek az agyalapi mirigybe. Hangsúlyozni kell, hogy a vazopresszin hormon prekurzora először a hipotalamuszban szintetizálódik. Ugyanakkor ott termelődik az 1-es és 2-es típusú neurofizin fehérje is. Az első az oxitocinhoz, a második a vazopresszinhez kötődik. Ezek a komplexek neuroszekréciós granulumok formájában vándorolnak a citoplazmában az axon mentén, és elérik az agyalapi mirigy hátsó lebenyét, ahol az idegrostok az érfalban végződnek, és a granulátumok tartalma a vérbe kerül. A vazopresszin és az oxitocin az első olyan agyalapi mirigy hormonok, amelyek teljesen kialakult aminosav-szekvenciával rendelkeznek. Kémiai szerkezetükben nonapeptidek, egyetlen diszulfidhíddal.
A vizsgált hormonok különféle biológiai hatásokat fejtenek ki: serkentik a víz és a sók transzportját a membránokon keresztül, vazopresszor hatásúak, fokozzák a méh simaizmainak összehúzódását a szülés során, és fokozzák az emlőmirigyek szekrécióját. Meg kell jegyezni, hogy a vazopresszin antidiuretikus aktivitása nagyobb, mint az oxitociné, míg utóbbi erősebb hatással van a méhre és az emlőmirigyre. A vazopresszin szekréciójának fő szabályozója a vízfogyasztás; a vesetubulusokban a citoplazmatikus membránok receptoraihoz kötődik, aminek következtében aktiválódik az azokban található adenilát-cikláz enzim. A molekula különböző részei felelősek a hormon receptorhoz való kötődéséért és a biológiai hatásért.
Az agyalapi mirigy, amely a hipotalamusz révén kapcsolódik az egész idegrendszerhez, funkcionális egésszé egyesíti az endokrin rendszert, részt vesz a szervezet belső környezetének állandóságának biztosításában (homeosztázis). Az endokrin rendszeren belül a homeosztatikus szabályozás az agyalapi mirigy elülső lebenye és a "cél" mirigyek (pajzsmirigy, mellékvesekéreg, ivarmirigyek) közötti visszacsatolás elve alapján történik. A "cél" mirigy által termelt hormon feleslege gátolja, hiánya pedig serkenti a megfelelő tropikus hormon szekrécióját és felszabadulását. A hipotalamusz a visszacsatolási rendszer része. Ebben helyezkednek el a "cél" mirigyek hormonjaira érzékeny receptorzónák. A vérben keringő hormonokhoz specifikusan kötődve és a hormonok koncentrációjától függően változtatva a választ, a hipotalamusz receptorai továbbítják hatásukat a megfelelő hipotalamusz központokba, amelyek koordinálják az adenohipofízis munkáját, hipotalamusz adenohipofiziotrop hormonokat szabadítva fel. Így a hipotalamuszt neuroendokrin agynak kell tekinteni.